






I cannot believe my good fortune to be a scientist
As told by Christopher Walsh
My earliest image of a scientist was a photograph of my older sister, Julie, on the wall of the attic bedroom that I shared with my two older brothers. She was captured by a school photographer while doing microbiology research at medical school, looking up from her Petri dishes and test tubes, hair pulled back behind her ear, pencil in hand, smiling. It looked like fun. As the seventh of eight children, I did not have much privacy growing up, but also little loneliness, sharing everything. Any spatial reasoning skills I have were honed in the kitchen, where whoever cut the pie got the last choice, so that slices were equivalent in size down to the gram. Even our dog had four names.
My father’s father was born in Pennsylvania – where the family settled from Ireland in the 1850s – but moved to Manhattan after medical school. There he became a prolific writer, especially focused on reconciling his Catholic faith with science and medicine, and married a schoolteacher. My father enlisted in the Army in World War II, and so had no time to train in medicine. With one child and a second on the way by war’s end, he chose the business world, commuting from our small bedroom town of Cranford, New Jersey into Manhattan very early every morning. He returned in time for dinner at a family table that was initially overflowing, but became progressively limited to my younger sister and me, as the older siblings left home.
My mother’s father was born in New York City, a few years after his parents’ arrival from Tvedestrand, Norway, working as a stonemason and raising a family in Brooklyn. My mother was an amazing person, a high school basketball star and a college graduate before her marriage, at a time when both were unusual for women. Raising eight children, she worked full-time while I was growing up to help support them through school, teaching math at high school and junior college. With her management and multitasking skills, she probably would have been a CEO in another era.


The Walsh family in the 1960s. Front row: Vincent, Arleen, Chris, Ellen, and Frances. Second row; James, Julie, Isabelle, and my parents Arleen Andersen Walsh and James Joseph Walsh.
Somehow, I was the only child who never attended Catholic school, instead taking a long bus-ride every day to Pingry, a rigorous independent school, for middle school and high school. There I discovered that I loved to learn. Unfortunately, the expensive school tuition forced a choice to preserve the family economy – three years of high school, or three years of college. And so, I found myself at Bucknell University, barely a month past my seventeenth birthday, with no idea of a college major or a life plan.
Two important courses
That uncertainty did not last very long, since my first semester featured two courses that have colored my work ever since. Whereas organic chemistry presented a beautiful spatial logic, psychology revealed that everything uniquely human, from language to art to music and consciousness, emerges from a brain that follows chemical rules. In many ways my subsequent career has bridged those two courses from that first semester.
“I remember one day seeing the chairman wagging a print back and forth in the air above his head to dry off the chemicals, while he gestured at us with that photo, saying “this will revolutionize neurology.”
My first lab job was at Columbia University, and that summer their first CT (computed tomography) brain scanner was installed. The grainy CT images were printed one-by-one as Polaroid images. I remember one day seeing the chairman wagging a print back and forth in the air above his head to dry off the chemicals, while he gestured at us with that photo, saying “this will revolutionize neurology.” At the time I had no idea what he was talking about, but it struck me that science could be really cool. On the other hand, I was undecided about medicine, because my experiences as a young volunteer were limited to work that lacked the magic of research. Entering the office of my pre-med advisor one day, I picked up a pamphlet from a pile near the door. That pamphlet described a program where one could do the M.D. and Ph.D. degrees together, in combined fashion, and that the NIH would provide full tuition and stipend support. To this day I will never forget that pamphlet – its burgundy ink, and its heavy, textured paper – and from then I was set on doing this combined degree program.
The University of Chicago was a perfect fit for me for graduate school, since it took “the life of the mind” very seriously, and I embraced that, finding time to read philosophy while taking medical courses. During my first year, my neighbor in the dormitory, an aspiring writer, developed a brain tumor, and my other friends and I watched his decline and death. I had not inherited my parents’ deep religious faith, so this experience challenged me spiritually in ways I still feel to this day, to find meaning and purpose in my life. It reinforced my desire to pursue science, as an opportunity to do something both creative and enduring, as well as to pursue medically related research so that my work might have impact on others.
Ray Guillery, my thesis advisor, became my role model for what a scientist could and should be. Ray could talk intelligently about seemingly anything, from opera to neuroscience to history, and as importantly he modeled that a scientist can be successful while being generous, collaborative, and also while raising a family, which I sensed would be important to me. Ray’s advice to me was to focus on a biological problem and use multiple methods to attack it: the problem I have been obsessed with ever since is how a collection of cells, with no special distinguishing features, somehow morphs into a human brain that transcends biology and psychology. Ray had discovered, among other things, that visual connections to the brain are systematically miswired in mammals (including humans) with albinism, the first example of genes regulating brain development in mammals. But perhaps more remarkable he found that higher visual centers, like the visual cortex, somehow recognize the abnormal wiring and suppress or even reorganize it, in a nongenetic fashion. This struck this young “armchair philosopher” as almost Kantian in its implications, as if our cortex regulates its own development, with an inherent demand for perception to be organized in a certain fashion. My own research with Ray, albeit more prosaic, analyzed the development of connections from eye to brain, to begin to understand how these ordered maps of the world get formed.


Ray Guillery and I on our way to the Society for Neuroscience conference in 2014. Ray had come through Boston from England, giving lectures at Harvard and MIT, and then received the Palay award at SFN. (picture by Kutay Deniz Atabay)
After graduating from Chicago, a residency in neurology at Massachusetts General Hospital (MGH) was the opportunity of a lifetime. The Chair of Neurology, Dr. Joseph Martin, had put the department at the cutting edge of using gene mapping to study human genetic neurological disorders like Huntington’s Disease, Alzheimer disease, and others. I’ll never forget Dr. Martin’s advice: “think about doing human genetics research, since it is the best of science, in the service of patients.” That advice resonated deeply with me, though the developmental disorders of human cerebral cortex that interested me were not feasible to study using gene mapping yet, never occurring in large families due to the severe disability they typically caused.
“I’ll never forget Dr. Martin’s advice: ‘think about doing human genetics research, since it is the best of science, in the service of patients’.”
The greatest gift
The greatest gift that MGH gave me was meeting my wife, Dr. Ming Hui Chen, a Harvard-educated cardiologist whom I met during my residency there. She has been my best friend and wisest advisor ever since, and we have raised two wonderful daughters together.
During and after residency I got the amazing opportunity to work with Dr. Connie Cepko in the Harvard Genetics department. Connie had developed the first way to map cell lineage in the brain, i.e., the formation of “families” of daughter cells that derive from a single brain stem cell. She invented viral vectors that label a stem cell, and all its daughter cells, with a marker that turns cells blue. Her studies in the retina showed that families of related cells formed tight-packed, almost crystalline blue arrays, so that a ray of light illuminates a clone of related retinal cells. I applied her method to the cerebral cortex, expecting to see similar blue clumps based on existing theory, but found instead that blue cells migrated away from one another, making it impossible to define where cells of one clone ended and the next clone began, since different clones all shared the same blue color. Typical of Connie’s fearlessness, when we discussed these results, her response was simple: “we just need a better technology.” So, I set about trying to make that better technology. By adding random DNA sequences inside the viruses – along with the blue color – the DNA barcodes, analyzed one cell at a time, would distinguish blue cells from different clones. Barcoded viral libraries showed that the cortex is a far more intermingled mosaic than the retina.


During and after residency I got the amazing opportunity to work with Dr. Connie Cepko in the Harvard Genetics department. Connie Cepko and I in 1992 trying to figure out the widespread patterns of clones in the cerebral cortex (Harvard photograph).
I have Dr. Clifford Saper to thank for my first independent faculty position, starting in 1993, at the Beth Israel Hospital, where he himself had just arrived as Chairman from the University of Chicago. As our new lab continued the study of cerebral cortical cell lineage in animals, I tried to find ways to study the mechanisms that guide these patterns of proliferation and migration.
A remarkable experience
A remarkable experience just months after starting my lab permanently changed the course of my research. At a meeting in Venice in May 1993 to present our data on cell lineage, I heard a presentation by Dr. Peter Huttenlocher, who had taught me pediatric neurology at Chicago. He described a family with an inherited cerebral cortical disorder, in which the neuronal migration that I was studying so intently completely failed to take place for some, but not all, of the neurons. He had tracked the disorder through the family by performing CT, or newer MRI imaging, to define those in the family who were affected. As his talk unfolded, I recall my heart racing and my palms sweating with excitement and anticipation, because here was the way that Joe Martin had suggested, and that I had been seeking, of doing the best science and improving medicine at once. Ming Hui was traveling with me, and we had lunch and a trip to Murano with Peter and his wife, Janellen (a noted psychologist herself), and planned our collaboration to find the gene for this condition. Amazingly, this particular condition, called periventricular nodular heterotopia (PVNH), is also associated with heart and vascular defects, so that Ming Hui and I have collaborated and published on it together ever since.


My first day as a human geneticist, in May 1993. I took this picture of Ming Hui with Janellen and Peter Huttenlocher while we ate lunch on Murano in Venice. That day the four of us planned our genetic collaboration to study PVNH.
No sooner did we set out to find the gene for Peter’s family than a second disorder of migration, double cortex syndrome presented itself as possible to map. My first postdoctoral fellow, Dr. Kristina Allen, mapped the double cortex condition, and she and Dr. Joseph Gleeson, a child neurologist, identified the gene for that condition, named doublecortin (DCX). That same year (1998) a student, Jeremy Fox, found FLNA mutations in PVNH. The striking finding in both conditions was that most affected patients did not inherit a mutation from parents, because the mutations are so debilitating that affected children rarely become parents. Instead, spontaneous mutations, occurring constantly with each generation, every now and then hit these genes to cause the conditions.
As we analyzed DCX and FLNA, finding dozens of families with mutations in each gene, I wondered whether these genes were unusual, or whether they were representative, which would mean that all human genes undergo the same onslaught of mutations. Testing this idea was inspired by another mentor, Dr. Verne Caviness, the Chief of Child Neurology at Massachusetts General Hospital, who had studied a mouse mutant named reeler, because of its reeling gait caused by a different type of neuronal migration disorder. The gene mutated in reeler, called Reelin, is one of the largest genes in our genome, so it seemed a likely target for mutation. Unlike DCX and FLNA that are X-linked, the reeler gene is autosomal recessive, so that affected mice carry two disabled copies of Reelin, inherited from unaffected parents, each with a single mutated Reelin gene and a second, normal version. It seemed that a human reeler disorder might be diagnosable by MRI like PVNH and double cortex, yet might occur in families where the parents are related as cousins: cousin parents are more likely to both be silent carriers of the same mutated gene, increasing the likelihood of recessive disease. To my surprise, there were several families described in the literature that looked like a human reeler disorder, and it took just two such families to show indeed that both families had Reelin mutations.
A whole new world of genetics
The Reelin work opened a whole new world of genetics to me. Recessive disorders are notoriously diverse, making them hard to study, yet families with shared ancestry offered the chance to find disease genes in very small families, and some families from which we already had DNA samples seemed likely to identify previously unidentified genes regulating cortical development. So, at this point (2000) I sent a FAX to every child neurologist I could find in the Middle East, where cousin marriages are common for important historical and cultural reasons, to invite collaborations on gene identification, and started traveling there to visit families and engage additional collaborations. These wonderful collaborations led, over the subsequent two decades, to identifying several dozen disease genes affecting all aspects of brain cell proliferation, survival, neuronal migration, and neuronal function.


Visiting a family with a recessive cognitive disorder in the mountains of Oman, ca 2009. On the left, Chiara Manzini, a postdoc at the time, climbs the mountain. On the right, she and Muna Al Saffar, our genetic counselor, and others take family histories and prepare to take blood samples.
As the lab expanded, I had the privilege to become a Howard Hughes Medical Institute (HHMI) Investigator in their first open competition in 2002. I would never have succeeded in that tough competition without the help of Clif Saper and Joe Martin, who had become dean of Harvard Medical School. The HHMI support challenged me to break new scientific ground, extending our Middle Eastern collaborations to study recessive causes of autism spectrum disorders. Autism is a genetically more complex condition, but one that promised to be more treatable than the structural malformations of the brain that had been our focus. The appeal of human genetics in this context is that it takes a problem of virtually infinite scope – the architecture of human behavior and cognition – and allows progressive, durable progress by referencing it to the genome. While the human brain evokes celestial comparisons in terms of its billions of neurons and their trillions of connections, the genome is finite, with about 20,000 genes, more like the inhabitants of a small town, and it is linear and digital in its primary organization.
The HHMI support also allowed me to re-design how I spent my time. Since 1993, I had maintained a general clinic in neurology, and spent one month each year on the inpatient neurology service at Beth Israel Deaconess Medical Center, work that was not related to our lab’s research program. With HHMI support, I focused specifically on the developmental disorders that we were studying in lab, starting a collaborative clinic with Boston Children’s Hospital (BCH) to diagnose and treat children with rare brain diseases. This program continues today as the BrDG (Brain Development and Genetics) program and has reinforced my sense of purpose, to see how the genetic insights that we gain in the lab can be transformative to families – even if we are not able to offer them a cure – and how the passion of families drives our research forward. Changing my clinical work also allowed me to be more involved in our daughters’ childhood. Our being able to drive together to and from school on most days until college made every day a good day.
“The BrDG (Brain Development and Genetics) program and has reinforced my sense of purpose, to see how the genetic insights that we gain in the lab can be transformative to families.”
Ideal home
Starting the BrDG program led to my recruitment to BCH in 2005, where I became Chief of the Division of Genetics. Dr. Gary Fleisher, the chief of pediatrics, has been an incredibly generous and visionary leader. His support provides a remarkable environment where my lab has been able to continue to be productive, while also helping build a division of amazing individuals, both scientists and clinicians, who care for children and adults with rare genetic diseases. Our division covers a range from diagnosis to the latest cutting-edge gene therapy trials for an ever-growing list of conditions. This has been the ideal home for our science.
As we identified genes essential for human brain development, we encountered a surprising subset of them whose DNA sequence systemically differed between humans and non-human species, apparently controlling aspects of cerebral cortical development that make the human brain unique. For example, several genes associated with microcephaly, characterized by a small cerebral cortex, show this evidence of positive evolutionary selection in humans, and these genes control when brain stem cells proliferate, and when they stop. This led to us finding that some parts of the genome showing the strongest evidence of evolution in humans are disrupted in patients with autism, suggesting essential roles for some of these sequences in human social behavior. This also led to funding from the Paul Allen Frontiers Program for a Center for Human Brain Evolution, along with Drs. David Reich and Mike Greenberg, to use human medical genetics to study human brain evolution systematically; since if those parts of the genome that are most uniquely human have roles in human brain, there will be disabilities associated with mutations that disrupt them, and hence can define their function.
Amidst our work on recessive disorders, the child neurologist Dr. Ann Poduri brought to my attention a condition that looked anything but inherited, because only half of the brain was abnormal, and the other half was normal. These children develop epilepsy that is so severe that the malformed parts of the brain are often removed surgically to treat the seizures, allowing us to study the abnormal tissue directly. We found that these patients had somatic mutations – mutations present in some cells but not all cells of the body – and mutations typically absent from blood DNA.
In order to assess how common these somatic mutations might be, two Ph.D. students, Gilad Evrony and Xuyu Cai, developed a method to amplify and analyze the entire genome of a single neuron and compare that genome to the one of the neuron next to it. This method showed that transposons could jump from one part of the genome to another during brain development, and that genomic segments could be duplicated or deleted during development as well. This analysis required a collaboration with Drs. Peter Park and Alice Lee, who have remained valued collaborators ever since.
A major turning point
A major turning point came when two postdoctoral fellows, Drs. Mike Lodato and Mollie Woodworth, analyzed these amplified single cell genomes in detail, finding thousands of potential mutations that distinguish each single neuron from the next. Some of these mutations form a developmental barcode, like those I had engineered as a postdoc, showing that each of us bears a permanent, forensic map of the development of our entire body. But the human developmental map is even more granular than we can create in animal models because every cell division in the embryo is marked by a handful of errors replicating the DNA of the genome, creating a series of clonal barcode mutations. Most of these developmental mutations are functionally silent, so are just lineage marks, but a subset of them, in some people, disrupt important genes that our lab has now shown create risk not just for focal epilepsy, but also for autism and schizophrenia. A major interest of the lab now is the role of this developmental map in generating the unique genetic endowment of each individual, and how somatic mutations create risk for other neurological diseases.
Surprisingly however, while some somatic mutations in single neurons represented barcodes shared between cells, a larger number of the mutations in the genome of a single neuron appeared to be unique to that neuron and not shared by other cells. How to deal with these? Were they just technical artifacts, or did a neuron really have a thousand unique mutations? How could a non-dividing neuron, that never replicates its genome, ever acquire so many mutations? We reasoned that if the neuronal genome was dynamically mutating, we should see biological patterns in the numbers and types of mutations at different ages, and in different brain areas, and in rare genetic forms of neurodegeneration. And, sure enough, we found that neurons are born with hundreds of somatic mutations, representing the developmental barcode; but then neurons continue to add mutations unique to a single cell, about 15-20 mutations per year, like a clock, so that neurons of elderly individuals have well over a thousand unique mutations. And recently, we found that this mutational process is not only accelerated in rare genetic conditions where neurons degenerate but is also accelerated in common forms of degeneration like Alzheimer’s disease. So, mutation accumulation is a marker of normal aging, and a marker – potentially a mediator – of age-related neuronal dysfunction, because when neurons have thousands of mutations, it becomes highly likely that essential genes are disrupted.
“But any success I have had reflects not my skill, but the intellect, creativity, and the dedication of those with whom I have had the privilege of working: postdoctoral fellows, PhD students, research assistants, and genetic counselors.”


The Lab celebrating at the neighborhood baseball park.
Good fortune
I cannot believe my good fortune to be a scientist, doing a job that is full of constant discovery. But any success I have had reflects not my skill, but the intellect, creativity, and the dedication of those with whom I have had the privilege of working: postdoctoral fellows, PhD students, research assistants, and genetic counselors. You can see that the early stages of my work bear the imprint of my teachers; but more recent work increasingly reflects my wonderful colleagues and trainees, who are also now my teachers of the newest and best science. I have also been fortunate to have experienced two remarkably warm and supportive Chairmen – Drs. Clifford Saper at Beth Israel and Gary Fleisher at Boston Children’s Hospital – who have helped our science in innumerable ways. But The Kavli Prize also recognizes that a field has reached a certain level of impact or maturity; in the case of the study of cerebral cortical development I am merely a representative of a wonderful community of scientists, men and women, who have inspired me by their own crucial contributions, and many of whom are as deserving as I to be singled out.
My own work has been deeply formed by my combined M.D.-Ph.D. training, and the discovery that every human – whether we like it or not – represents an experiment of nature in some way and provides a way to discover science that we could not otherwise have imagined. It has been a privilege to be a part of the Harvard-MIT combined M.D.-Ph.D. training program, and to have spent several years as the director of that program. Our science has also been indelibly shaped by the influence of HHMI, which has repeatedly funded new directions that have been highly risky but have shaped our science. Other funders, especially the NINDS, NIMH and the Allen Frontiers Program, have also been essential to our success. But finally, I come back again to family, where it begins. I have been incredibly fortunate to have siblings who get along well to this day, and we all miss our parents, whom we lost more than a decade ago now. And my success would mean nothing if I could not share it with Ming Hui, and to have been able to have such a wonderful family of our own.

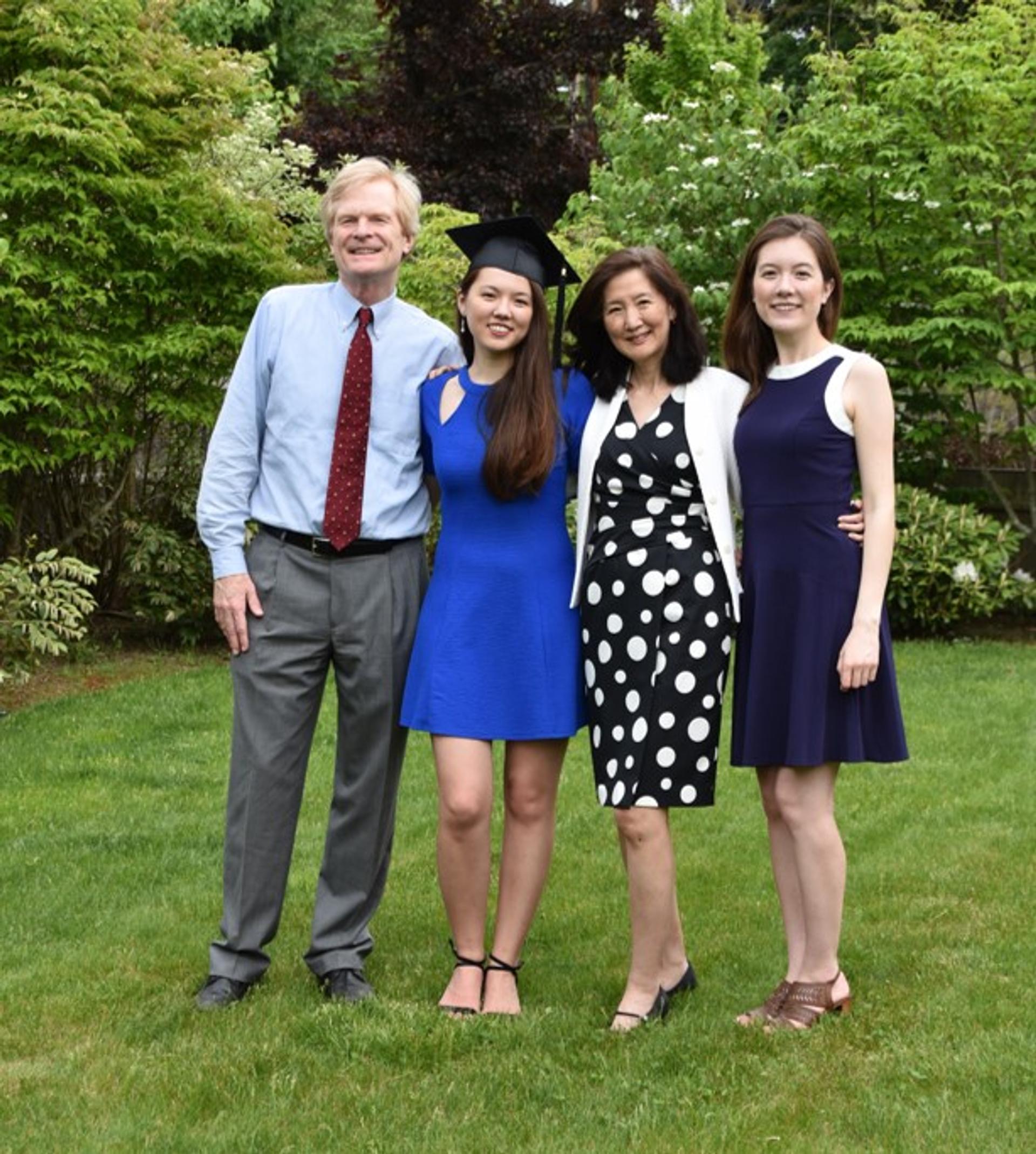
The Chen-Walsh family in 2020, celebrating our younger daughter’s backyard graduation from college during the pandemic, with her older sister. (picture courtesy of Stephanie Scanlon).